

Biological interactions are necessary for the functioning of an ecosystem and also for the survival of the individual organism as no organism can exist in absolute isolation in nature. Look at our surroundings, climbers twining around the trees for support, orchids and mosses grow on moisture laden branches, Cuscuta or golden parasites spread over shrubs/trees, all are the intricate relationship between host and seekers in terms of habitat, nutrition, and protection. Symbiosis is one such interrelation in which organisms are involved in a close and long-term relationship with each other and each termed a symbiont and may be of the same or of different species. Mutualism is a type of symbiosis in which all the participating organisms benefit from the relationship. We come across mutualism every day around us in nature, for example when we see a tree log covered in lichen inside a forest which is actually a complex symbiotic association between algae or cyanobacteria and fungi. The relationship of lichen is mutualistic in nature as both the species get benefits i.e. fungi helps in anchorage, protection and absorbing nutrients from the substrate while the algae contribute in photosynthesis by using their chlorophyll that the fungi lack. The similar mutualistic symbiotic association we also see in mycorrhiza fungi of Glomeromycota as they form an association with the root system of higher plants and contribute extensively to nutrients exchange.
Myrmecophytes or ant-plants are an excellent example of a symbiotic relationship where the plant has a symbiotic relationship with ant colonies. Symbiotic ant-plants often develop a hollow structure in their stem internodes called a domatium that provides shelter to its ant resident and also provides the food resources in the form of extrafloral nectar (EFN). In return, most associated ant species protect the plant against herbivores, pathogens and other competing vegetation (Frederickson et al., 2005; Rosumek et al., 2009).
Humboldtia brunonis (Leguminosae), an endemic ant-plant of the Indian Western Ghats dominates the rainforest understory (Figure 1) and produces EFN on young expanding leaves and floral bud bracts but all individual plants do not produce domatia (Chanam et al., 2015). Plants with domatia give shelter to 16 ant species of which only Technomyrmex albipes provides protection to the plant in the region where herbivory is highest and Vombisidris humboldticola is uniquely associated with the plant (Zacharias & Rajan, 2004; Shenoy & Borges, 2010; Shenoy et al., 2012; Chanam et al., 2014). The domatia are also occupied by several other invertebrates, most prominently by the arboreal earthworm Perionyx pullus (Gaume et al., 2006) (Figure 2). Domatia-bearing plants receive more benefits in terms of nitrogen absorbed from ant-occupied domatia (Chanam et al., 2014).
the Indian Western Ghats dominates the rainforest understory (Figure 1) and produces EFN on young expanding leaves and floral bud bracts but all individual plants do not produce domatia (Chanam et al., 2015). Plants with domatia give shelter to 16 ant species of which only Technomyrmex albipes provides protection to the plant in the region where herbivory is highest and Vombisidris humboldticola is uniquely associated with the plant (Zacharias & Rajan, 2004; Shenoy & Borges, 2010; Shenoy et al., 2012; Chanam et al., 2014). The domatia are also occupied by several other invertebrates, most prominently by the arboreal earthworm Perionyx pullus (Gaume et al., 2006) (Figure 2). Domatia-bearing plants receive more benefits in terms of nitrogen absorbed from ant-occupied domatia (Chanam et al., 2014).
Ant-plant systems are often tripartite or even more complex where third-party interactions are provided by ant-associated mutualistic fungi (Mayer et al, 2014). Ant–fungal mutualisms are 45–65 million years old and widely known in fungus-farming attine ants (subfamily Myrmicinae, tribe Attini) where the ants cultivate fungi as the food source. Many ants use fungi to build nests, traps, or galleries using carton structures (cardboard-like materials from masticated plant fibers) (Lauth et al., 2011).
The fungal culture is also reported in few groups of arboreal ants dwelling inside domatia of tropical ant-plants where the ants are reported to maintain a group of fungi inside domatia as a possible food source (Blatrix et al., 2009; Vasse et al., 2017). Domatia-dwelling ants also use fungi to construct carton structures in order to modify the domatia interiors in certain systems as well (Baker et al., 2017; Chanam and Borges, 2017).
We have found that in Humboldtia brunonis system ants use fungi in two distinctive ways inside the domatia: i) building a disc-shaped carton partition between the zone occupied by ants and earthworms in the case of co-occupied domatia, and ii) maintaining a group of fungi in the inner wall of domatia, probably as a food source (Figure 3). We hypothesized that in the presence of earthworms, with which the ants have an antagonistic relationship, the ants can only reside and maintain a fungal culture in the domatia wall by constructing a carton separation using a different group of fungi. 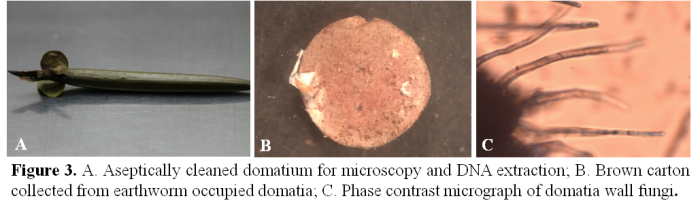 To disentangle the fungal association with ant-plant complex, we approached through metagenomics to characterize two distinct fungal communities directly from domatia sample as many fungi still non-culturable in laboratory condition. We characterized the community profile of car-ton and wall fungal clades by the metabarcoding technique in the DNA sequencing platform named Oxford Nanopore using universal fungal genomic marker ITS1 (Figure 4). When we successfully identify some organism we call them a taxon but in many cases using genomic marker especially for enigmatic microbes, we cannot identify the exact taxon name. In a scenario like this, we use a terminology called Operational Taxonomic Unit or OTU. We have found that the OTU abundance for both fungal clades indicates the ants have a clear preference for fungi of the order Chaetothyriales as a probable food source in the domatia wall while they use a more diverse group of fungi in cartons. Our study shows a possible tripartite symbiosis in this ant–plant system involving ants, plants, and fungi for the first time from the Indian subcontinent.
To disentangle the fungal association with ant-plant complex, we approached through metagenomics to characterize two distinct fungal communities directly from domatia sample as many fungi still non-culturable in laboratory condition. We characterized the community profile of car-ton and wall fungal clades by the metabarcoding technique in the DNA sequencing platform named Oxford Nanopore using universal fungal genomic marker ITS1 (Figure 4). When we successfully identify some organism we call them a taxon but in many cases using genomic marker especially for enigmatic microbes, we cannot identify the exact taxon name. In a scenario like this, we use a terminology called Operational Taxonomic Unit or OTU. We have found that the OTU abundance for both fungal clades indicates the ants have a clear preference for fungi of the order Chaetothyriales as a probable food source in the domatia wall while they use a more diverse group of fungi in cartons. Our study shows a possible tripartite symbiosis in this ant–plant system involving ants, plants, and fungi for the first time from the Indian subcontinent.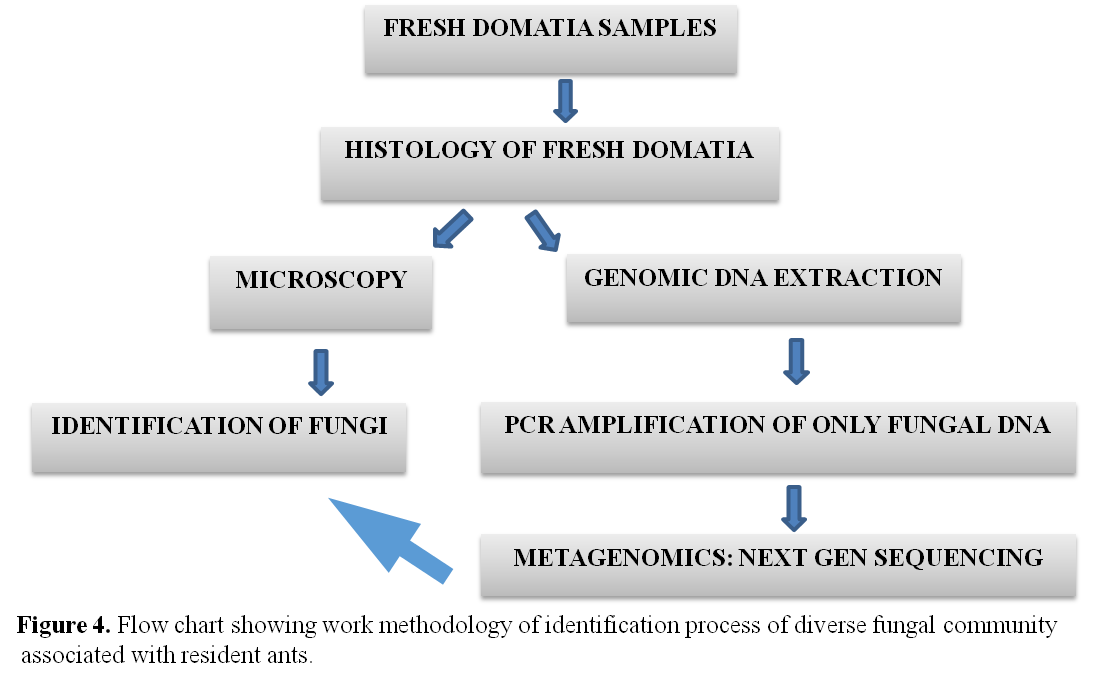
Fungi have been noticed within domatia of other ant-plants (Janzen, 1972). It has been demonstrated in the symbiosis between the plant-ant Petalomyrmex phylax and the ant-plant Leonardoxa africana (Defossez et al., 2009) that the ants provide protection and nutrients to their fungal symbiont (Defossez et al., 2011) and use the fungal symbiont as a food source (Blatrix et al., 2012). Another function of fungi associated with plant-ants may be to enhance the transfer of nutrients to the host plant (Leroy et al., 2011). Fungi are also reported to be manipulated by ants to make net-like traps for capturing prey (Nepel et al., 2014). So far the fungi known to be involved in the tripartite ant–plant–fungus interactions belong to the order Chaetothyriales (Ascomycota). These fungi are usually dark, melanized, slow-growing, often colonize extreme environments (Zhao et al., 2010; Seyedmousavi et al., 2011) and are commonly called ‘‘black yeasts’’ (Voglmayr et al., 2011).
Our findings have showed similar fungal preference by ant but in this system, the ants are using the fungi in both as a probable food source and building block elements in presence of other invertebrates i.e. the earthworms. Most interestingly the ants are perfectly aware of the specific fungal groups which are supposed to be used for food and which are supposed to be used for the building block.
Mutualism has its contribution to the fitness benefits and also in nitrogen budget of the plant either directly through absorption by the plant from ant wastes or via fungi cultivated by the ants within the domatia (Rosumek et al. 2009; Defossez et al. 2011; Leroy et al. 2011; Dejean et al. 2012, 2013). Similarly, in Humboldtia system, trees with domatia has greater fruit set than those without domatia; if the domatia were occupied by the protective ants then the fitness benefit was found to be even greater (Gaume et al. 2005). The Humboldtia system provides a unique opportunity to explore further the behavioral aspects of ants towards a specific group of fungi in the presence or absence of different co-occupants. The exact purpose of fungi and any other role of their presence that might felicitate benefits to the plants still remain to be explored. How the ants are able to maintain the different group of fungi in close domatia is a surprising fact and yet to be discovered in this fascinating endemic symbiotic ant plant system from the Indian Western Ghats.
References:
1) Baker CCM, Martins DJ, Pelaez JN, Billen JPJ, Pringle A, Frederickson ME & Pierce NE (2017) Distinctive fungal communities in an obligate African ant-plant mutualism. Proceedings of the Royal Society London B, 284: 20162501.
2) Blatrix R, Bouamer S, Morand S & Selosse MA (2009) Ant-plant mutualisms should be viewed as symbiotic communities. Plant Signaling and Behaviour, 4: 554–556.
3) Blatrix R, Djiéto-Lordon C, Mondolot L, Fisca PL, Voglmayr H & McKey D (2012) Plant-ants use symbiotic fungi as a food source new insight into the nutritional ecology of ant-plant interactions. Proceedings of the Royal Society London B, 279: 3940– 3947.
4) Chanam J & Borges RM (2017) Cauline domatia of the ant-plant Humboldtia brunonis (Fabaceae). Flora, 236-237: 58– 66.
5) Chanam J, Kasinathan S, Pramanik GK, Jagdeesh A, Joshi KA & Borges RM (2014) Context dependency of rewards and services in an Indian ant-plant interaction: southern sites favour the mutualism between plants and ants. Journal of Tropical Ecology, 30: 219-229.
6) Chanam J, Kasinathan S, Pramanik GK, Jagdeesh A, Joshi KA & Borges RM (2015) Foliar extrafloral nectar of Humboldtia brunonis Fabaceae, a Paleotropic ant-plant is richer than phloem sap and more attractive than honeydew. Biotropica, 47: 1-5.
7) Defossez E, Djiéto-Lordon C, McKey D, Selosse MA & Blatrix R (2011) Plant-ants feed their host plant but above all a fungal symbiont to recycle nitrogen. Proceedings of the Royal Society London B, 278: 1419-1426.
8) Defossez E, Selosse MA, Dubois MP, Mondolot L, Faccio A, Djieto-Lordon C, McKey D & Blatrix R (2009) Ant-plants and fungi a new three-way symbiosis. New Phytologist, 182: 942-949.
9) Dejean A, Petitclerc F, Roux O, Orivel J & Leroy C (2012) Does exogenic food benefit both partners in an ant-plant mutualism? The case of Cecropia obtusa and its guest Azteca plant-ants. Comptes Rendus Biologies, 335: 214–219.
10) Dejean A, Orivel J, Rossi V, Roux O, Lauth J, Male PJG. et al. (2013) Predation success by a plant-ant indirectly favours the growth and fitness of its host myrmecophyte. PLoS ONE, 8: e59405.
11) Frederickson ME, Greene MJ & Gordon DM (2005) Ecology: ‘Devil’s gardens’ bedeviled by ants. Nature, 437 : 495-496.
12) Gaume L, Zacharias M, Grosbois V & Borges RM (2005) The fitness consequences of bearing domatia and having the right ant partner: experiments with protective and non-protective ants in a semi-myrmecophyte. Oecologia, 145: 76–86.
13) Gaume L, Shenoy M, Zacharias M & Borges RM (2006) Co-existence of ants and an arboreal earthworm in a myrmecophyte of the Indian Western Ghats: anti-predation effect of the earthworm mucus. Journal of Tropical Ecology, 22: 1–4.
14) Janzen DH (1972) Protection of Barteria (Passifloraceae) by Pachysima ants (Pseudomyrmecinae) in a Nigerian rain forest. Ecological Applications, 53 : 885-892.
15) Lauth J, Ruiz-González MX & Orivel J (2011) New findings in insect fungiculture: Have ants developed non-food, agricultural products?. Communicative & Integrative Biology, 4(6): 728–730.
16) Leroy C, Séjalon-Delmas N, Jauneau A, Ruiz-González MX, Gryta H, Jargeat P, Corbara B, Dejean A & Orivel J (2011) Trophic mediation by a fungus in an ant-plant mutualism. Journal of Ecology, 99: 583-590.
17) Mayer VE, Frederickson ME, McKey D & Blatrix R (2014) Current issues in the evolutionary ecology of ant-plant symbioses. New Phytologist, 202: 749–764.
18) Nepel M, Voglmayr H, Schonenberger J & Mayer VE (2014) High diversity and low specificity of chaetothyrialean fungi in carton galleries in a neotropical ant–plant association. Plos One. 9(11): 1-10.
19) Rosumek FB, Silveira FAO, Neves FDS, Barbosa NPDU, Diniz L, Oki Y, Pezzini F, Fernandes GW & Cornelissen T (2009) Ants on plants: a meta-analysis of the role of ants as plant biotic defences. Oecologia, 160: 537-549.
20) Seyedmousavi S, Badali H, Chlebicki A, Zhao J, Prenafeta-boldú FX & Hoog GSD (2011) Exophiala sideris, a novel black yeast isolated from environments polluted with toxic alkyl benzenes and arsenic. Fungal Biology, 115: 1030-1037.
21) Shenoy M & Borges RM (2010) Geographical variation in an ant-plant interaction correlates with domatia occupancy local ant diversity and interlopers. Biological Journal of the Linnean Society, 100: 538-551.
22) Shenoy M, Radhika V, Satish S & Borges RM (2012) Composition of extrafloral nectar influences interactions between the myrmecophyte Humboldtia brunonis and its ant Associates. Journal of Chemical Ecology, 38: 88-99.
23) Vasse M, Voglmayr H, Mayer VE, Gueidan C, Nepel M, Moreno L, de Hoog S, Selosse MA, McKey D & Blatrix R (2017) A fungal phylogenetic perspective of the association between ants (Hymenoptera: Formicidae) and black yeasts (Ascomycota: Chaetothyriales). Proceedings of the Royal Society London B, 284: 20162519.
24) Voglmayr H, Mayer V, Maschwitz U, Moog J, Djieto-Lordon C & Blatrix R (2011) The diversity of ant-associated black yeasts: insights into a newly discovered world of symbiotic interactions. Fungal Biology, 115: 1077-1091.
25) Zacharias M & Rajan PD (2004) Vombisidris humboldticola (Hymenoptera: Formicidae): a new arboreal ant species from an Indian ant plant. Current Science, 87: 1337–1338.
26) Zhao J, Zeng J, Hoog GSD, Attili-Angelis D & Prenafeta-Boldú FX (2010) Isolation and identification of black yeasts by enrichment on atmospheres of monoaromatic hydrocarbons. Microbial Ecology, 60: 149-156.
About Author :


